Effect of pharmacological concentrations of zinc oxide with or without the inclusion of an antibacterial agent on nursery pig performance
Journal of Animal Science, January 2001
A study involving nine research stations from the NCR-42 Swine Nutrition Committee used a total of 1,978 crossbred pigs to evaluate the effects of dietary ZnO concentrations with or without an antibacterial agent on postweaning pig performance. In Exp. 1, seven stations (IA, MI, MN, MO, NE, ND, and OH) evaluated the efficacy of ZnO when fed to nursery pigs at 0, 500, 1,000, 2,000, or 3,000 mg Zn/kg for a 28-d postweaning period.
A randomized complete block experiment was conducted in 24 replicates using a total of 1,060 pigs. Pigs were bled at the 28-d period and plasma was analyzed for Zn and Cu. Because two stations weaned pigs at < 15 d (six replicates) and five stations at > 20 d (18 replicates) of age, the two sets of data were analyzed separately. The early-weaned pig group had greater (P < 0.05) gains, feed intakes, and gain:feed ratios for the 28-d postweaning period as dietary ZnO concentration increased. Later-weaned pigs also had increased (P < 0.01) gains and feed intakes as the dietary ZnO concentration increased. Responses for both weanling pig groups seemed to reach a plateau at 2,000 mg Zn/kg. Plasma Zn concentrations quadratically increased (P < 0.01) and plasma Cu concentrations quadratically decreased (P < 0.01) when ZnO concentrations were > 1,000 mg Zn/kg. Experiment 2 was conducted at seven stations (KY, MI, MO, NE, ND, OH, and OK) and evaluated the efficacy of an antibacterial agent (carbadox) in combination with added ZnO. The experiment was a 2 × 3 factorial arrangement in a randomized complete block design conducted in a total of 20 replicates. Carbadox was added at 0 or 55 mg/kg diet, and ZnO was added at 0, 1,500, or 3,000 mg Zn/ kg. A total of 918 pigs were weaned at an average 19.7 d of age. For the 28-d postweaning period, gains (P < 0.01), feed intakes (P < 0.05), and gain:feed ratios (P < 0.05) increased when dietary ZnO concentrations increased and when carbadox was added. These responses occurred in an additive manner. The results of these studies suggest that supplemental ZnO at 1,500 to 2,000 mg Zn/kg Zn improved postweaning pig performance, and its combination with an antibacterial agent resulted in additional performance improvements.
Introduction
Nursery studies evaluating the efficacy of pharmacological concentrations of dietary Zn on postweaning pig performance have generally demonstrated positive growth responses (Holm, 1988, 1990; Poulsen, 1989; Hahn and Baker, 1993). These responses have been achieved at dietary concentrations of 2,000 to 4,000 mg Zn/kg (Poulsen, 1995; Smith et al., 1997; Hill et al., 2000). The oxide form of the mineral seems to be critical in achieving these response benefits and is less toxic than other inorganic Zn sources (Hahn and Baker, 1993; McCully et al., 1995; Schell and Kornegay, 1996). High dietary concentrations of ZnO increased liver Zn (Schell and Kornegay, 1996; Carlson et al., 1999) but decreased liver Fe concentrations (Cox and Hale, 1962). Carlson et al. (1999), however, did not affect liver Cu or Fe concentrations when high dietary ZnO concentrations were fed to weanling pigs for a 2- to 4-wk period.
Copper sulfate provided at dietary concentrations substantially higher (125 to 250 mg Cu/kg) than the NRC (1998) requirement (6 mg Cu/kg) has also resulted in improved nursery pig performance (Braude, 1967; Prince et al., 1979; Edmonds et al., 1986). The data from Beames and Lloyd (1965) demonstrated an additive growth response when a high dietary Cu concentration was fed in combination with an antibiotic. Studies using various antibiotics or antibacterial agents in combination with CuSO4 have also demonstrated an additive performance effect with weanling pigs (Stahly et al., 1980; Roof and Mahan, 1982). When high concentrations of CuSO4 (250 mg Cu/kg) or ZnO (2,000 or 3,000 mg Zn/kg) were tested, growth responses occurred with each mineral source but were not additive (Smith et al., 1997; Hill et al., 2000). Experiments were conducted by the North Central Regional Swine Nutrition Committee (NCR-42) to further evaluate the 1) feeding of various dietary concentrations of ZnO on postweaning pig performance and blood Zn and Cu concentrations and 2) feeding of an antibacterial agent in combination with ZnO on weanling pig performance.
Materials and Methods
Experiment 1
The first experiment evaluated the efficacy of five dietary concentrations of supplemental ZnO fed to postweaning pigs in a randomized complete block design. The concentrations evaluated were 0, 500, 1,000, 2,000, and 3,000 mg Zn/kg using a common source of ZnO that contained 72% Zn (Prince Agri Products, Quincy, IL). Seven university research stations (IA, MI, MN, MO, NE, ND, and OH) participated in the experiment for a total of 24 replicates (each contributed a minimum of two replicates). The experiment included 1,060 crossbred pigs of various genetic crosses that were weaned at an average age of 19.1 d (range 11 to 25 d) and an average BW of 6.0 kg (range 3.9 to 8.4 kg). Upon weaning, pigs were allotted to treatment pens on the basis of weight, sex, and litter using an equal number of pigs per pen within each replicate. Each station differed in its facility, management, and laboratory methodology, but the procedures followed within each replicate were identical. Pigs were housed in nursery facilities that contained either slotted concrete or plasticcovered wire, mesh-floored pens, with room temperatures adjusted when needed to meet the comfort zone of the pig. Pig information and other experimental conditions for the seven stations that conducted the experiment are presented in Table 1.
Postweaning diets contained both animal and vegetable protein sources and were formulated to provide a dietary lysine concentration of 1.35% (total) for the initial 14-d postweaning period and 1.15% (total) lysine for the 14- to 28-d postweaning period. Vitamin and trace mineral premix compositions differed among stations but each premix met or exceeded NRC (1988) requirements. Each station added the antibiotic chlortetracycline at 220 mg/kg to the nursery diets. Supplemental ZnO was added to treatment diets at the appropriate concentration at the expense of corn. The composition of the basal diets is presented in Table 2. Diets were subsampled at mixing and subsequently analyzed for Zn, Fe, and Cu. The analyzed basal diets averaged 291 and 300 mg Zn/kg, 265 and 280 mg Fe/kg, and 16 and 15 mg Cu/kg for the two production phases, respectively.
Pigs consumed their treatment diets on an ad libitum basis in meal form during the 28-d experimental period. Pig weights and feed intakes were collected weekly. At the end of the 28-d study, blood was collected from the vena cava from all or from randomly selected pigs within each treatment pen; a similar number of pigs was bled in each pen for each replicate. Pigs from six replicates (n = 350 pigs) in the early-weaned group and 14 replicates (n = 480 pigs) in the later-weaned group were bled, and the blood was centrifuged and the plasma separated, frozen, and subsequently analyzed for Cu and Zn.
Experiment 2.
Because of the positive growth response to added ZnO in Exp. 1, this experiment was conducted to evaluate whether there was an interaction between ZnO and an antibacterial agent. The experiment was a 2 × 3 factorial arrangement of treatments and was conducted in a randomized complete block design. Carbadox (Mecadox; Pfizer Animal Health, Terre Haute, IN) inclusion (0 or 55 mg/kg) was the first factor and the dietary concentration of ZnO (0, 1,500, or 3,000 mg Zn/kg) was the second factor tested. The experiment was conducted at seven university research stations (KY, MI, MO, NE, ND, OH, and OK). Each station contributed a minimum of two replicates for a total of 20 replicates. A total of 918 crossbred pigs of differing genetic crosses were weaned at an average age of 19.7 d (range 14 to 21 d) and an average BW of 6.5 kg (range 5.8 to 7.0 kg). Although cooperating stations were not identical to those of Exp. 1, experimental conditions were generally similar to those in the previous experiment and identical within replicate. Pig and experimental facility information for the cooperating stations is presented in Table 1.
Basal diets contained various dietary protein sources for each production phase; their compositions are presented in Table 2. The diets for the initial week postweaning contained 1.50% (total) lysine, followed by a 1.30% (total) lysine concentration during the subsequent 7 to 28-d period. The composition of the vitamin and trace mineral premixes differed among stations but all met or exceeded NRC (1988) standards. Dietary ZnO was added to treatment diets at the appropriate concentration at the expense of corn. Diets were sampled at the time of mixing and subsequently analyzed for Zn. The basal diets analyzed an average of 142 and 194 mg Zn/kg for the two phases, respectively. Diets were provided in meal form and the pigs were allowed to consume the diets on an ad libitum basis. Performance responses were evaluated for the 0- to 7- and 7- to 28- d periods.
Analytical Methods
Each station followed the approved experimental and bleeding procedures of their respective animal care committees. Subsamples of treatment diets and the individual plasma samples were sent to Michigan State University for analysis. Diet samples of Exp. 1 were ground, digested in nitric and perchloric acid, and analyzed for Fe, Zn, and Cu as outlined by Hill et al. (1983), whereas the treatment diets of Exp. 2 were analyzed only for Zn. Plasma samples were diluted 1:7 with deionized water, and Zn and Cu concentrations were determined by flame absorption spectrophotometry (Smith-Hieftje 4000, Thermo Jarrell Ash Corp., Franklin, MA). Standards for Fe, Zn, and Cu obtained from the National Institute of Standards and Technology were used for instrument standardization and quality control.
The data of both experiments were statistically analyzed using the analysis of variance procedures of Steel and Torrie (1980) and the GLM procedures of SAS (SAS Inst. Inc., Cary, NC). For the statistical models of both experiments the treatment, station, and replicate effects were considered fixed. In Exp. 1, two stations had weaned their pigs at < 15 d of age, whereas five stations weaned their pigs at > 20 d. Consequently, the performance and blood mineral data from these two station groups were analyzed and reported separately. The effect of ZnO concentration for all measurement variables in both experiments were contrasted by regression analysis. Pen averages were considered the experimental unit for all traits in both experiments.
Results
Experiment 1
The performance data partitioned into the early (11 to 15 d) and later (21 to 25 d) weaning age groupings are reported in Tables 3 and 4, respectively. Pigs weaned at the earlier age did not have an increased gain or feed intake response to dietary ZnO concentrations during the initial week postweaning. However, during the following 7- to 14-d period the gain responses to dietary ZnO concentrations were linear (P < 0.01), whereas during the 14- to 21-d and 21- to 28- d periods the gains increased in a quadratic manner (P < 0.05). Feed intake during the 7- to 14-d and 14- to 21-d periods increased linearly (P < 0.01) to added ZnO, and from 21 to 28 d the response was quadratic (P < 0.01). Gain:feed ratio also increased linearly (P < 0.01) for the 7- to 14-d period and quadratically (P < 0.05) during the 14- to 21-d period, but there was no significant treatment effect during the 21- to 28-d period. For the overall 28-d period pig gains and gain:feed ratio showed a quadratic (P < 0.05) increase to ZnO concentrations. These responses seemed to indicate that the plateau occurred when the 2,000 mg Zn/kg concentration was provided. When the 28-d performance response of pigs fed 2,000 mg Zn/kg was compared to that of pigs fed the basal diet, gains increased an average of 29%, feed intake 20%, and the gain:feed ratio 7%.

In contrast to the earlier weaned group, the pigs that were weaned at the older age (21 to 25 d) had increased gains, feed intakes, and gain:feed ratios as the dietary ZnO concentration increased during the initial week postweaning for each measurement variable (P < 0.01). During the 7- to 14-d and the 14- to 21-d periods gains increased (P < 0.01) as the dietary ZnO concentration increased, but the gain response for the 21- to 28-d period was not significant (P > 0.15). Feed intakes were greater at each weekly measurement period as dietary ZnO concentration increased, but treatment differences were not significant (P > 0.15) for the 7- to 28-d period. Gain:feed ratios increased as dietary ZnO concentration increased at each weekly measurement period and were significant (P < 0.01) for the 14- to 21-d period. The response was quadratic (P < 0.01) for the overall 28-d period. As with the early-weaned group, performance responses seemed to plateau at the 2,000 mg Zn/kg concentration. For the overall 28-d period, pig gains, feed intakes, and gain:feed ratios increased by approximately 10, 5, and 5%, respectively, when the pigs fed the 2,000 mg Zn/kg diet were compared to those that were fed the basal diets. Plasma Cu and Zn concentrations at the end of the 28-d postweaning period responded in an inverse manner to each other as dietary ZnO increased for both weaning age groups (Table 5). Plasma Cu decreased quadratically (P < 0.01) whereas plasma Zn increased quadratically (P < 0.01) as dietary ZnO increased; the greatest change occurred between the 2,000 to 3,000 mg Zn/kg concentration.
Experiment 2
From 0 to 7 d postweaning there was no gain response to dietary ZnO when carbadox was added to the diet, but there was a positive (17%) growth response to ZnO concentrations when the diet did not contain carbadox (Table 6). This resulted in a carbadox × ZnO concentration interaction response (P < 0.05) for the initial week postweaning. During the 7- to 28-d period (P < 0.01) and for the overall 28-d postweaning period (P < 0.05), daily gains increased when carbadox was added to the diet, with quadratic increases (P < 0.01) to added ZnO. There was no interaction (P > 0.15) between the two treatment variables during either measurement period, indicating that the responses to ZnO and carbadox were additive.
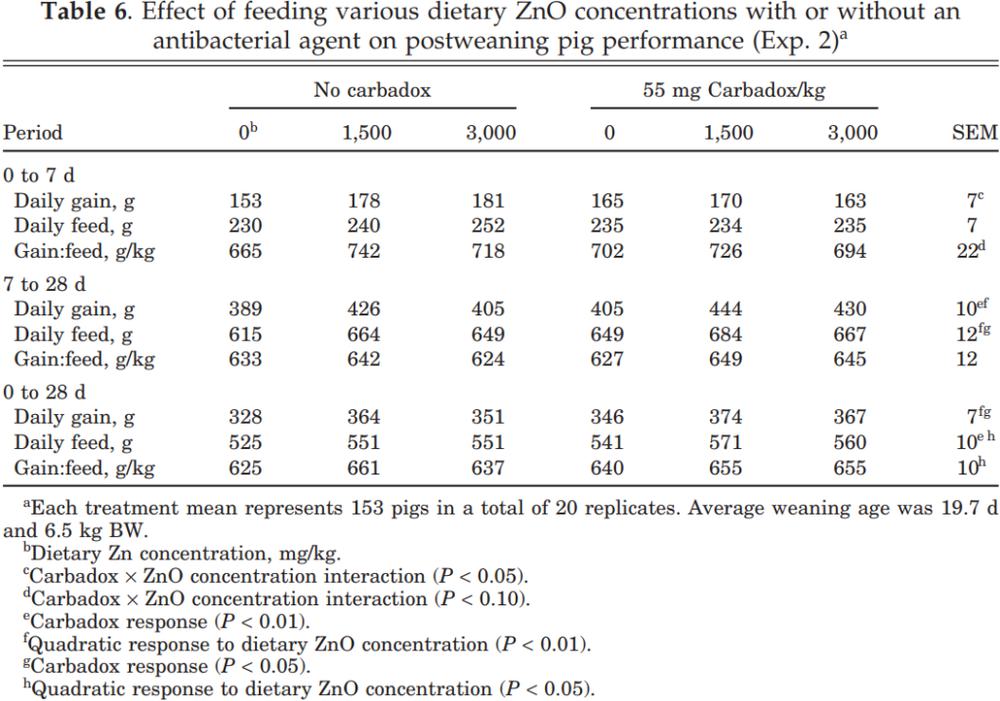
Feed intake from 0 to 7 d postweaning was not affected by carbadox or by ZnO concentration (P > 0.15). For the 7- to 28-d period, feed intake increased when carbadox was added to the diet (P < 0.05) with a quadratic increase (P < 0.01) in feed intake as the ZnO concentration increased. For the overall 28-d period daily gains and feed intakes both increased (P < 0.01) when carbadox was added, with quadratic improvements (P < 0.01, P < 0.05, respectively) for both variables as the ZnO concentration increased. Gain:feed ratios tended (P < 0.10) to improve during the 0- to 7-d period when both ZnO and carbadox were added to the diet but quadratically (P < 0.05) when the ZnO concentration increased. For the 0- to 28-d period, these measurements seemed to reach a plateau at the 1,500 mg Zn/ kg concentration, and the responses were generally consistent with those of Exp. 1.
Discussion
Much of the early work that evaluated high dietary concentrations of ZnO centered on the hypothesis that this inorganic mineral source reduced postweaning pig mortality and diarrhea that was attributable to E. coli, whereupon pig gains increased (Holm, 1988, Poulsen 1989). Subsequent research with weanling pigs in the United States demonstrated increased pig growth responses when high concentrations of ZnO were fed to weanling pigs without conditions of diarrhea (Hahn and Baker, 1993; Carlson et al., 1999; Hill et al., 2000). Consequently, ZnO is now generally included in many nursery pig diets at dietary concentrations of > 3,000 mg Zn/kg as a growth promotant and as an aid in preventing postweaning diarrhea.
Our experiment evaluated the efficacy of dietary ZnO at nine different university research stations using a total of 1,978 pigs that involved various genotypes, facilities, management conditions, and weaning ages. The results clearly demonstrated a growth response to added ZnO that reached a plateau at dietary concentrations of 1,500 to 2,000 mg Zn/kg.
When pigs were weaned early (< 15 d of age) the growth response to 2,000 mg Zn/kg averaged 29% for the 28-d feeding period, whereas the growth response with later-weaned pigs was somewhat lower, averaging 10%. In the early-weaned pig group, no growth response to ZnO occurred during the 1st wk postweaning, but increased growth, feed intake, and feed efficiency responses occurred during the subsequent 21-d period. At those stations where pigs were weaned at 20 d of age or later, most of the growth response occurred during the initial 21-d postweaning period, with a positive but smaller response during the 21- to 28-d period. The reason for the differing time responses to supplemental ZnO between the two weaning ages is unclear. Carlson et al. (1999) reported that both early- and later-weaned pigs also responded in a beneficial manner to supplemental ZnO from weaning to 14 d postweaning with minimal differences in responses thereafter.
The efficacy of supplemental carbadox on postweaning pig performance responses has been well established (Thrasher et al., 1969; Yen et al., 1976; Roof and Mahan, 1982). In our study the response to ZnO seemed to reach a plateau at 1,500 mg Zn/kg and was independent of the added carbadox. Our data thus indicated that the addition of the antibacterial agent (i.e., carbadox) and ZnO each resulted in an independent growth response, but when added together the responses were additive. Whether this effect would occur with other antibacterial agents is not known.
Although the basal diets contained an average of 295 mg Zn/kg in Exp. 1 and 168 mg Zn/kg in Exp. 2 (both in excess of NRC [1998] requirements), the performance responses achieved from ZnO inclusion at all stations occurred only when a high supplemental concentration of ZnO was provided. Consequently, even with the wide range of Zn concentrations in the various basal diets (range of 47 to 506 mg Zn/kg) the addition of ZnO at dietary concentrations of ≥1,500 mg Zn/kg was essential to achieve improved nursery pig performances. Environmental and management conditions that existed in the various nursery facilities differed, but growth responses to the combination of ZnO and carbadox during the 28-d postweaning period occurred at each station and averaged 10% in Exp. 2.
The increase in plasma Zn and the concurrent decline in plasma Cu began when dietary Zn concentrations were > 1,000 mg/kg and was exacerbated as the dietary Zn concentration increased. This effect occurred in both early- and later-weaned pigs and suggests that the plasma homeostasis of Zn and Cu might be disturbed when dietary Zn concentrations are > 1,000 ppm. Hahn and Baker (1993) and Carlson et al. (1999) demonstrated that plasma Zn concentration increased as the dietary concentration of Zn increased, particularly when diets contained > 1,000 mg Zn/kg. The form of Zn in the diet can, however, affect its bioavailability and subsequent plasma Zn concentration (Hahn and Baker, 1993). These latter workers reported that ZnO had a lower bioavailability, which resulted in lower plasma Zn concentrations compared to Zn sources chemically bound with sulfate, carbonate, or the ligands of lysine or methionine. Cox and Hale (1962) demonstrated that when high Zn concentrations were fed liver Fe concentrations were reduced and anemia developed. Carlson et al. (1999) reported that although metallothionein in the intestinal tract and liver increased when high-ZnO diets were fed, no effect on liver Fe or Cu concentrations occurred when high concentrations of ZnO were fed for relatively short periods postweaning.
Our results therefore suggest no benefit when supplementing nursery pig diets with ZnO at dietary concentrations in excess of 1,500 to 2,000 mg Zn/kg. Because many commercial feed companies are incorporating higher dietary concentrations of Zn that could disturb the body’s homeostasis of these or other elements, this continued practice is questionable. With the low bioavailability of Zn from ZnO much of the Zn is excreted in the feces. Consequently, lower dietary concentrations of ZnO (1,500 to 2,000 mg Zn/kg) when fed for relatively short periods (≤ 28 d) would seem to pose fewer environmental concerns than the current practice of supplementing higher concentrations of the mineral. #
Implications
Adding ZnO to nursery pig diets resulted in increased pig performance responses that reached a plateau at dietary concentrations of 1,500 to 2,000 mg Zn/kg. The performance responses to ZnO and the antibacterial agent carbadox were additive. Earlier-weaned pigs may benefit more from high dietary ZnO concentrations, but a positive growth and feed response also occurred in later-weaned pigs.
