Modern genetics has resulted in chickens characterised by high performance. The downside of this success is the high sensitivity to numerous stresses. To allow the animal the chance to respond adequately to these stresses, selenium (Se) is added to the diet. As an essential trace element, selenium is required for optimal performance, reproduction and immunity. Specific enzymes (selenoproteins) require the incorporation of Se to ensure their activity in the animal. These enzymes reduce the presence of reactive oxygen species, produced during normal metabolic activity and are elevated during stress causing cell damage and subsequent malfunction of tissues. Protection against these harmful metabolites is especially important in high energy-demanding cells.
The new divide: L-selenomethionine compared to all other selenium compounds
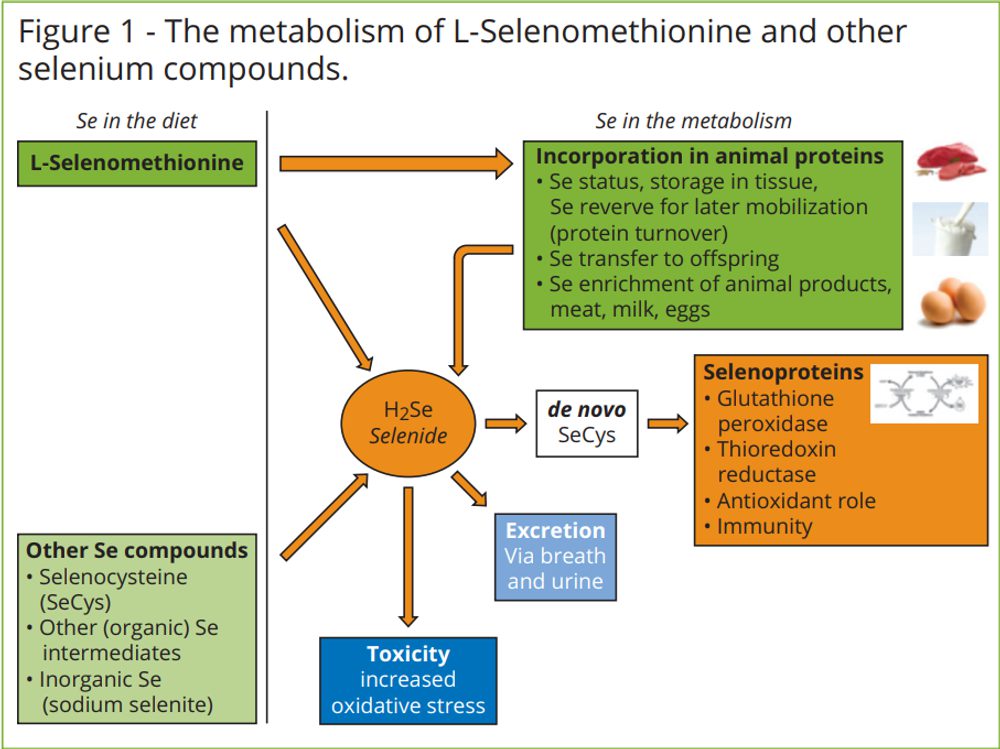
Se can be added to the diet as L-selenomethionine or as sodium selenite, sodium selenate, selenocysteine or organic selenium intermediates. The advantage of selenium in the form of Lselenomethionine (L-SeMet) over all other selenium compounds is the ability to be incorporated directly into animal proteins. This metabolically active Se acts as a storage depot for Se within tissues (Figure 1). Other Se compounds are invariably reduced to hydrogen selenide (H2Se) and subsequently converted to de novo selenocysteine and incorporated into selenoproteins. Hydrogen selenide does not function as a reserve for Se in tissues and cells and any excess is excreted via methylation in the liver to prevent toxic reactions.
L-SeMet, is the only Se compound that can be directly, without conversion, incorporated into structural proteins in muscle and liver tissue. This storage ensures optimal Se supply, even during stress such as heat exposure when dietary intake of Se can be limited. Under stress conditions there is a high demand for Se to inactivate oxygen free radicals and reduce cell damage. Research has shown that Se from sodium selenite is excreted at a rate almost three-fold more than Se from L-SeMet when fed to broilers, confirming that inorganic Se is not stored (Skrivan et al. 2008).
L-SeMet is generally supplied in diets by addition of selenized yeast. Yeasts have limited ability to incorporate Se in the form of L-SeMet, (Schrauzer, 2006). Normally, around 63% of the total Se in a selenized yeast is present as L-SeMet but there is a large variation among products and batches (Figure 2). L-SeMet incorporated in yeast protein is also not completely bioavailable as it has to be digested before it can be absorbed. (Sauvant,2004). Other selenium sources present in yeast cells follow the pathway to hydrogen selenide and are therefore not any more valuable as a Se source than inexpensive inorganic Se. From an efficiency point of view, selenized yeast does not provide the full potential of L-selenomethionine. Pure synthetic preparations of L-selenomethionine, contributing selenium as a single amino acid is the most bioavailable form.
The benefits of L-selenomethionine in poultry
Frequent stressful periods are always correlated with an increased metabolic activity and energy demand that is correlated with a period of high oxidative stress. To combat this stress the animal requires proportionately higher levels of Se. The literature confirms that heat- stressed chick embryos benefit from maternal Se in the form of L-SeMet compared to inorganic Se (Xiao et al. 2016). A significant increase in selenoprotein synthesis including glutathione peroxidase occurs with a resulting reduction in reactive oxygen species (ROS) and malondialdehyde, a biomarker for lipid peroxidation. L-SeMet is beneficial in improving the antioxidant response of chick embryos as it is transferred efficiently and to a high extent from the diet to the egg. When comparing sodium selenite, selenized yeast and L-SeMet in deposition of Se in the egg, the L-SeMet was able to increase the Se content in the egg to the highest level among alternative sources (Delezie et al. 2014).
L-SeMet is not only able to adequately respond to heat stress in embryos, but also in broilers as it the compound is stored efficiently in muscle tissue as a component of muscle proteins. When broilers were subjected to chronic heat stress during the finisher period, feeding L-SeMet improved weight gain and enhanced the efficiency of feed conversion (Michiels et al. 2016).
After slaughter L-SeMet has a positive effect on meat quality. Chick tissue is especially vulnerable to oxidative stress expressed as lipid peroxidation due to the presence of long-chain polyunsaturated fatty acids (PUFA). L-SeMet not only reduces lipid peroxidation but also increases the alpha-tocopherol (vitamin E) content due to inherent regenerative ability that is more efficient than sodium selenite (Skrivan et al. 2008). Studies have demonstrated a reduction in drip loss of almost one percent after storage of the meat for 48 hours when diets were supplemented with L-SeMet compared to sodium selenite (Zhang et al. 2014).