A disruption of this steady state causes tissue damage due to interaction of ROS with lipids, proteins and DNA. These negative interactions reduce their metabolic activity.
In order to maintain this steady state a continuous as well as optimal selenium supply is essential. However, this can be difficult to achieve when uptake from the diet is impaired when stress is present. At that moment selenium is in high demand, to produce selenoenzymes and combat ROS. Selenium storage inside the animal, in that respect, would be beneficial. This article provides an overview of the scientific literature on the beneficial effects seen with the addition of L-selenomethionine to the diet with focus on salmon, trout and tilapia. Results from a recent trial on tilapia conducted in Thailand are discussed.
Maintaining an optimal selenium steady state: A nutritional solution
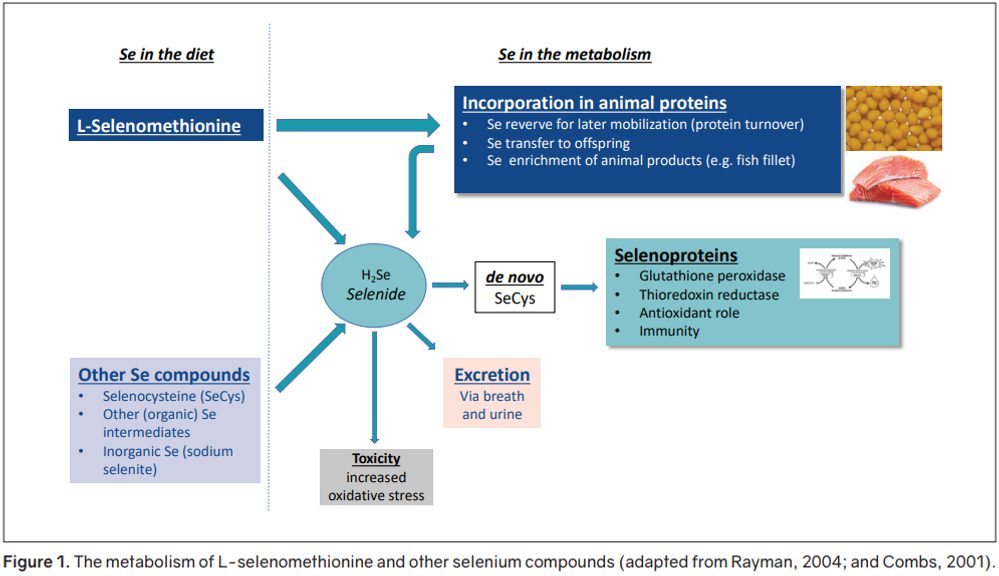
Selenium can be added to the diet in either inorganic or organic forms (Figure 1). The advantage of using organic selenium (L-selenomethionine, L-SeMet) over inorganic sources (e.g. sodium selenite or selenate) is its ability to be incorporated directly, without conversion, into general body proteins as a methionine source. L-selenomethionine is the only selenium compound that has this ability. The incorporated selenium, in the form of L-selenomethionine, acts as a storage of selenium in the animal. This stored selenium ensures optimal supply, even during stressful periods.
If necessary, the stored selenium gets metabolised to selenide (H2Se) then to de novo selenocysteine (SeCys). This molecule will be incorporated, as the active site, in selenoproteins. Other selenium compounds, such as SeCys and sodium selenite, are not storable but will be metabolised to de novo SeCys. These compounds will be quickly excreted when intake is in excess. L-selenomethionine will only be metabolised to selenide when there is a need. This form is therefore less prone to excretion and toxicity reactions (Rayman, 2004).
Aquatic protein challenge: A case for L-selenomethionine
Traditionally, fishmeal is the preferred protein source in aquatic feeds. Due to limited availability, pressure on wild fish stocks and variable prices, there is an interest in alternative, sustainable protein sources. Plant meals, for example, are suitable alternatives in the growing global aquaculture industry. However, replacing marine ingredients in fish feed with plant sources changes the nutrient composition of the feed. Selenium concentration of fillets is reported to be highly impacted by high levels of substitution, reducing the added value of fish consumption (Lundebye et al. 2017; Betancor et al. 2016).