Its development is directly or indirectly associated with increased risk of other conditions such as: cardiorespiratory, orthopedic alterations and metabolic disorders such as reduced glucose tolerance, type 2 diabetes in felines, hyperlipidemia (Gayet et al., 2004; Brunetto et al., 2011) and reduced life expectancy (Kealy et al., 2002).
The mechanism involved in the development of many of the mentioned alterations relies on the characterization of adipocytes as members of an active endocrine tissue (Balistreri et al., 2010). Therefore, adipose tissue also participates in mechanisms of inflammation and immunity, with production and release of a wide range of pro-inflammatory factors such as interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α), and suppression of anti-inflammatory factors such as adiponectin, as well as other cytokines.
Weight loss is able to reduce these changes, the severity of complications, and even normalize the alterations acquired by overweight status. The success of the treatment depends on the promotion of weight loss and effective maintenance of lean weight, thus it is necessary to induce a negative energy balance state by means of caloric restriction (Cloetens et al., 2012; Floerchinger et al., 2015). For such, satiety plays a major role during this process concerning the animal and its owner, since the manifestation of hunger, which consequently leads the animal to seek and beg for food, often compromises the continuation of the program (Weber et al., 2007).
Beta-1,3/1,6-glucans are polysaccharides composed of glucose monomers linked together by β-glycosidic bonds. Due to their complex mechanism of action in the organism, several effects have already been associated with their supplementation: modulation of the immune response (Li et al., 2006; Zaine, 2014), reduction of the inflammatory response (Beynen and Legerstee, 2010; Beynen et al., 2011; Silva et al., 2015) and modulation of glucose (Vetvicka and Oliveira, 2014; Silva et al., 2015) and lipids (Andrade et al., 2016) metabolism. However, the effects of beta-1,3/1,6-glucans on glycemic control are poorly understood in dogs, and no studies that evaluated the addition of this polysaccharide in glycemic parameters of obese dogs are available.
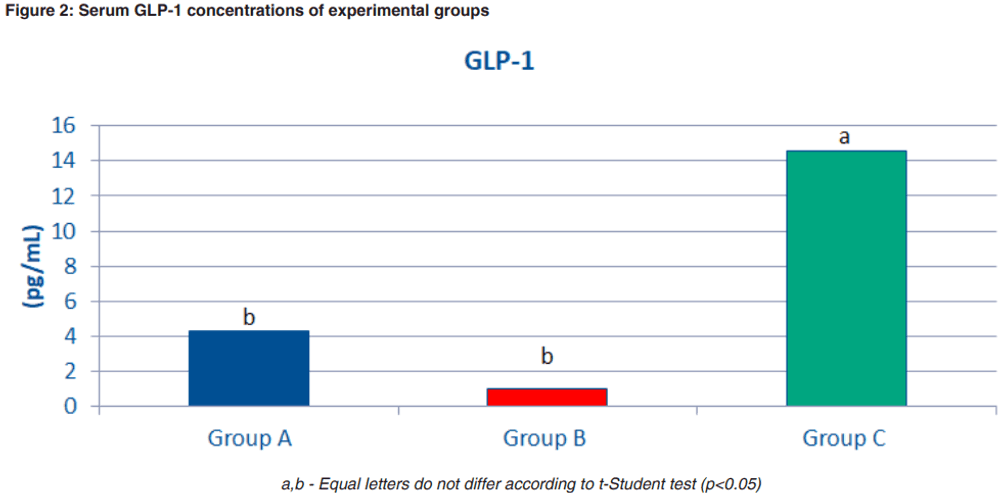
A recent study carried out at the School of Veterinary Medicine and Animal Science – University of São Paulo (Brazil) evaluated the effects of purified beta-1,3/1,6-glucans from yeast S. cerevisiae (MacroGard, Biorigin, Brazil) on metabolic parameters and hormones related to satiety of obese dogs. Fourteen privately owned dogs were split into two experimental groups: obese dogs (group A) with body condition score (BCS) 8 or 9/9 and lean dogs with BCS 5/9 (group B), according to the Laflamme (1997) scoring system. For a period of two weeks, both groups received a control diet (no beta-1,3/1,6- glucans). After two weeks, group A became group C, and received a diet with beta-1,3/1,6-glucans (MacroGard®) for a period of 90 days. The trial set up is shown in Figure 1.