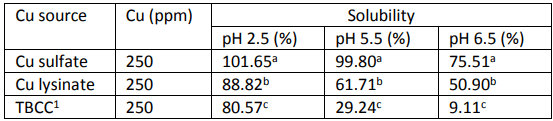
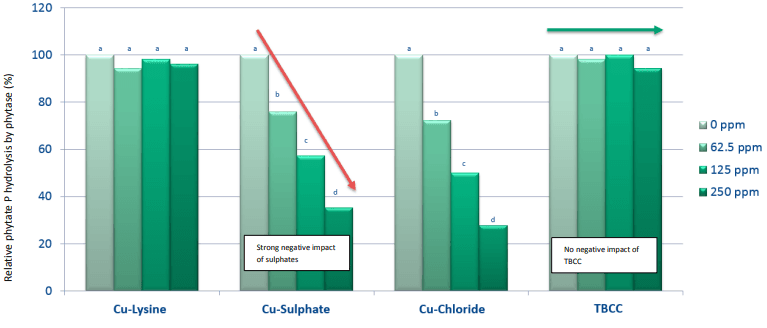
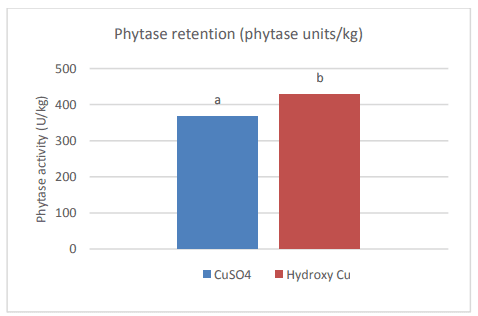
Graph 2: Effect of Cu sulphate and hydroxy Cu chloride on phytase retention. (a, b)Values with different letters are different (P < 0.05) from each other.
Next to Phytase, soluble Cu also has a negative effect on the fat content in feed. Dissociated, soluble Cu can initiate the lipid autoxidation cycle by playing a role as catalysator in the formation of free radicals, hereby reducing the nutritional value of the fat. A large difference was shown between the oxidation capacity of Cu sulfate and hydroxy Cu chloride (Miles et al., 1998). In this study, Feed grade Cu sulfate was sieved and coarse and fine Cu sulphates were retained with a respective modal particle size of 455 µm a b 0 100 200 300 400 500 Phytase activity (U/kg) Phytase retention (phytase units/kg) CuSO4 Hydroxy Cu Strong negative impact of sulphates No negative impact of TBCC Relative phytate P hydrolysis by phytase (%) and 11,9 µm. The two Cu sulphate fractions and hydroxy Cu chloride (modal particle size = 67 µm) were mixed in broiler diets at 25 ppm Cu. The peak peroxidation value observed for fine Cu sulphate was 5,8 times higher than for hydroxy Cu sulphate. At day 20, the anisidine value measured for fine Cu sulphate was 2,2 times higher than for hydroxy Cu sulphate. The values for coarse Cu sulphate were comparable to hydroxy Cu chloride at all time points and for both measured parameters. When using feed grade Cu sulphate, there will always be a part of fine size and this part will have a larger negative effect on lipid peroxidation compared to hydroxy Cu chloride. Although the average particle size of hydroxy Cu chloride was smaller than coarse Cu sulphate, the effect of oxidation was similar. Improved oxidative stability of soybean oil towards hydroxy trace minerals compared to sulphates was confirmed in an in vitro model using Rancimat Test (Kampf et al., 2012).
To assess the slow release effect of hydroxy Cu chloride throughout the GIT an in vivo study was conducted (Naziripour & Klasing, 2010). Broilers were fed 150 ppm Cu, from Cu sulphate and hydroxy Cu chloride, for 2 weeks (day 3 – 14). The amount of Cu (mg/kg DM) was analyzed and divided into water soluble Cu, EHPG extractable Cu and unremovable Cu (unavailable for nutritional and microbicidal purposes). Copper sulphate was shown to increase the duodenal luminal soluble Cu almost twofold compared to hydroxy Cu chloride, augmenting the negative effect of Cu on the villi and the tendency to form complexes with dietary nutrients resulting in a higher unremovable Cu part later on in the GIT. Hydroxy Cu chloride resulted in more EHGP-extractable Cu in all regions of the intestines (P < 0.05) and less unremovable Cu (P = 0.05), relating to its greater bioavailability and anti-coli activity. Hydroxy Cu chloride was shown to inhibit E.coli growth (P < 0.05) to a greater extent than Cu sulphate in the lower gastrointestinal tract. Performance-wise there is also a clear distinction between Cu sulphate and hydroxy Cu chloride (Kim & Kil, 2015) The inclusion of 100, 200 and 300 mg/kg added Cu from hydroxy Cu chloride resulted in an improved ADG, ADFI and FCR for all levels compared to Cu sulphate.
Hydroxy Zinc chloride has been researched in several animal species. Relative bio-availability was measured in piglets where improved bio-availability was shown for hydroxy zinc compared to zinc sulphate ranging from 122% (metacarpal zinc) to 159% (plasma zinc after 2 weeks of supplementation) (Zhang and Guo, 2007). In broilers, inclusion of 80 ppm hydroxy zinc chloride resulted in heavier birds and more breast meat yield compared to 80 ppm of zinc sulphate or a combination of 60 ppm zinc sulphate combined with 20 ppm zinc-chelate, without compromising feed conversion (Parr et al., 2013)
Selenium in feed – all about L-selenomethionine
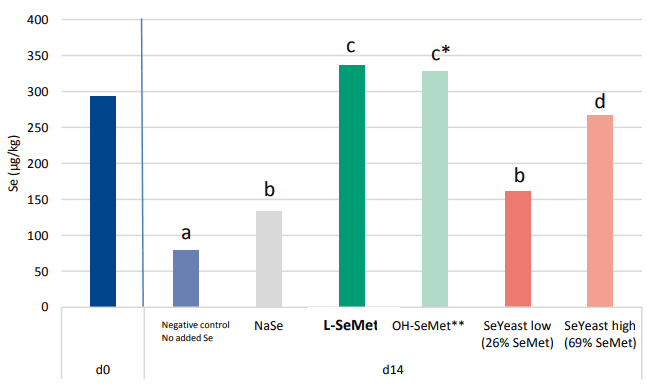
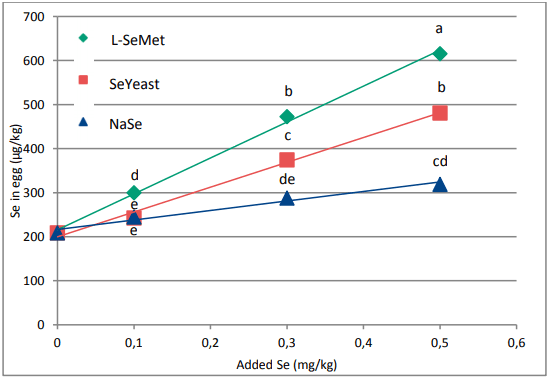
When looking at selenium, different sources are used in practice. While selenite was and still is used for a big extend in feeds, it is generally accepted that organic selenium is the best source of selenium. Organic selenium sources are evaluated on their content of L-Selenomethionine (L-SeMet). This unique form can be build up, in the same way as methionine, in the protein fraction of tissues and animal products (milk, eggs, meat). A study proved the enhanced Se deposition of L-SeMet compared to other selenium containing (in)organic sources in broiler breast meat (Graph 3; Van Beirendonck et al., 2016). Two Seyeasts were incorporated in the trial reflecting products that can be found in the market. The Se-yeast high in Se as L-SeMet (69% of the total Se content) showed a high deposition but significantly lower than the group fed L-SeMet. The Se-yeast low in L-SeMet (26%) showed even a significantly lower deposition compared to the Se-yeast high in L-SeMet. When plotting the Se deposition in function of the L-SeMet concentration, it can be clearly derived that the deposition is linearly proportional to the L-SeMet concentration in the product. OH-SeMet has a numerical lower deposition of Se in the muscle compared to L-SeMet. When the non-intended overdose is taken into account, the selenium deposition confirmsthe efficacy of 80% for the conversion of OH-SeMet to L-SeMet in the animal, as described for OH-Methionine (EFSA Journal 2012;10(3):2623).